| Contents | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
MoleculeMolecules interact with each other to build pathways. A molecule is anything that is subject to reactions. Most molecules have a mass, be it a small molecule like ATP, or a protein. No difference is made between receptors, enzymes, second messengers, transcription factors or other special kinds of proteins. A molecule can also be a group of such entities, like a protein family, a state of such an entity, like the phosphorylated form or a complex of several other molecules. And finally a molecule can be part of another molecule, either non-covalently bound as in a complex, or covalently bound as in a structural motif of a protein. The reason for such a wide scope for this class is to catch anything that shows specific signaling behavior. A molecule can serve in four fundamental roles in the context of a reaction: it can be an educt, a product, an enzyme or a modulator. Enzyme, educt and modulator are the inputs of the reactions, while products are the outputs. For semantic reactions molecules are only grouped into signal donors, which are the inputs, and signal acceptors, which are the outputs. We use two other collections, modified forms and complexes, to keep the states/modified forms and complexes separated from the family grouping hierarchy.
FamilyGroupingImportant attributes of all components are the collections that allow expression of family relationships. Each component can act as a group of other components or as a member in such a group. In the case of molecule, this grouping represents family relationships between the molecules. In the case of reaction, this grouping can be used to assign reactions to a general class of reactions, as it is done successfully in the EC nomenclature. The third possibility is the use of mixed groups, which contain both reactions and molecules, and correspond to nets or explicitly stated pathways. This is interesting since it would support the appealing idea of building networks from subnets in a modular fashion.
Grouping molecules and motifs into families is essential for a usable signal transduction database. When the family relationships are stated in a formal way, it is easy to write algorithms that exploit them in user queries, while marking inherited properties as inferred information. Family specifically addresses the issues of
The family relation is implemented by the groups/members collections.
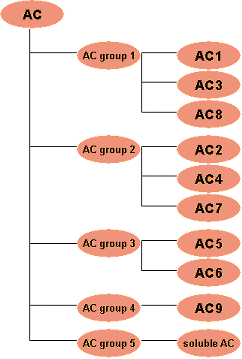
IsoformsDue to gene duplication and mutation within species, multiple paralogues have often evolved. For a single gene, different splice variants may exist. Sometimes in the literature a signaling activity is first attributed to a single molecule, and later it is discovered that there is a whole group of similar molecules. Therefore, a special type of molecule entry is needed, which we term "isoform". This abstracted group entry collects all the non-specific information. For example, look at adenylate cyclase (AC, Fig. 1). There are at least ten isoforms (AC1-9, sAC), which are all activated by Gsa, while only isoforms AC1, AC3 and AC8 are Ca/CaM dependent, AC2, AC4 and AC7 have no inhibition mechanism and AC5 and AC6 are inhibited by Gia or calcium.
OrthologsIn many cases, reactions between proteins have only been shown for one of several model species, such as for rat, clawed frog or fruit fly. It is common to scan sequence databases for ortholog sequences and assume, if these can be found, that they play a similar role in signal transduction. This orthologous grouping makes it possible to stitch together pathways from bits and pieces that have been investigated in different species.
Classification methodsSince it is easier to examine the sequence of a gene or protein than to investigate its function, a certain behavior is usually shown in detail for only few members of a protein family, and homologues are added to the functional group by sequence or structural similarity. Based on this assumption, various good databases exist that try to classify proteins and map sequence motifs to functional annotation. They cluster proteins by multiple sequence alignments and use common structural motifs, "profile" patterns, or Hidden Markov Models derived from these alignments to classify new proteins. Sometimes these methods can correctly predict the function, but sometimes not. Thus it is common practice to group molecules into families on the basis of sequence similarities, even if they do not share common behavior. For a signaling database, it is advantageous to group molecules that show common signaling behavior, that is, to group them by function. Since it is the function we are interested in, we would like to group only by function. On the other hand, we would like to draw as much as possible upon expert knowledge, and stay coherent with the groupings that already exist. To solve this dilemma, the best option seems to be that we group the molecules as it is done traditionally, but link signaling only to those molecules for which it has been shown. Given the tree for adenylate cyclase in Fig. 1, if the calmodulin-dependent reaction has been demonstrated for ACI, we link that reaction to ACI only. We link statements made on a generalized level, like the ones from review papers, to nodes on a higher level in the molecule hierarchy. Given the tree we can, for example, link the general Gsa activation to the AC group. The context of the original literature statement is preserved.
States (modified forms)The unmodified form of a protein and all its modified forms are its states, where the modification can be by covalent binding, by complexation, or by change of environment. The protein per se is a concept that is based on the observation that there is only one gene coding for each protein sequence. All the states share the same gene and consequently part of their structure, the amino acid chain. They are functionally related, often even reversibly transformable into each other. A basic molecule entry captures this concept and is the class of all states of a protein. These states are different molecules, and we store them as different molecule entries. As molecules, they can be used in a pathway assembly. We store general information like the amino acid sequence in the basic molecule entry and link its states to it. In the simplest case there are only two states- an inactive one, and an active one. In other cases, there are more. For example, a transcription factor can be
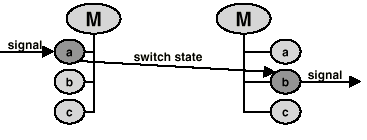
to name a few possibilities. The same protein will exhibit distinctly different signaling functions in these three states. For example in state 1 it will be susceptible to phosphorylation, in state 2 to translocation into the nucleus or dephosphorylation and in state 3 it will activate transcription. The number of states for a molecule is the product of the number of its modified forms and the number of locations where it is found. Only compounds which share the same location interact in nature. It is impractical to enter a separate state for each location. Most molecules can be found in several tissues, at several development stages, in several cellular compartments, several organs and several celltypes. To enter a state for each possible combination would lead to an explosion in the number of states, and redundancy in the reactions. This problem is circumvented by using a list of positive and negative locations that is linked to the basic molecule. In each state, the molecule is available only for a subset of all reactions for that molecule. Receiving a signal changes the molecule's state, usually leading to a new state from which reactions are triggered (Fig.2).
Motifs are structural and functional features of a protein and are often responsible for its signaling behavior. A single protein can have several signaling motifs. Motifs are listed in the feature field together with position information and references.
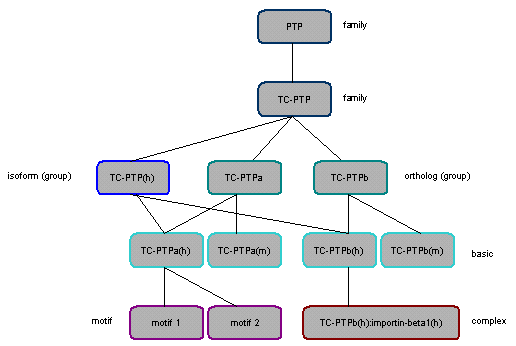
Figure 3 sums up all the various hierarchical relations that molecules in the database can have.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||